|
Arctic and Antarctica
Reference:
Vasil'chuk, A.C., Vasil'chuk, Y.K. (2021). Verification of paleo reconstructions of climatic conditions of the Late Pleistocene summer seasons based on palynological analysis of Yedoma deposits and ice wedges. Arctic and Antarctica, 4, 15–31. https://doi.org/10.7256/2453-8922.2021.4.36787
Verification of paleo reconstructions of climatic conditions of the Late Pleistocene summer seasons based on palynological analysis of Yedoma deposits and ice wedges
Vasil'chuk Alla Constantinovna
Doctor of Geography
Leading Research Fellow, Laboratory of Geoecology of the Northern Territories, Faculty of Geography, Lomonosov Moscow State University
119991, Russia, g. Moscow, Leninskie Gory, GSP-1, 1,, geograficheskii fakul'tet, NIL geoekologii Severa

|
alla-vasilch@yandex.ru
|
|
 |
Other publications by this author
|
|
|
Vasil'chuk Yurii Kirillovich
ORCID: 0000-0001-5847-5568
Doctor of Geology and Mineralogy
Professor, the department of Geochemistry of Landscapes and Geography of Soils, M. V. Lomonosov Moscow State University
119991, Russia, G Moscow, oblast', g. G Moscow,, ul. Leninskie Gory, 1, of. 2009
|
|
alla-vasilch@yandex.ru
|
|
 |
|
DOI: 10.7256/2453-8922.2021.4.36787
Received:
05-11-2021
Published:
27-01-2022
Abstract:
The object of this research is the reconstruction of the climatic conditions of the Late Pleistocene summer seasons based on the pollen spectra of syngenetic ice wedges and inclosing Yedoma deposits. For quantitative assessment of summer temperatures and phenological relationships, the author applies the total annual thaw index, since namely the sum of positive temperatures is the only indicator that determines the beginning of pollen emission in majority of plant species. The relationship between the composition of pollen spectra and the total annual thaw index is deterministic, although it cannot be formulated strictly mathematically. Yedoma is a unique object for research by many parameters, which allows acquiring paleogeographic information on two components, such as ice wedges and deposits, because the accumulation of pollen and spores occurs differently therein. The structure of vegetation cover during the accumulation of Yedoma deposits during the Late Pleistocene differed significantly from the Holocene and modern periods, which is recorded in the pollen spectra of syngenetic deposits and ice wedges. It is established that in certain periods, the diversity of species of Late Pleistocene vegetation was higher compared to modern vegetation of tundra and forest tundra due to the large number of ecological niches and peculiarity of temperature regime of the summer season.
Keywords:
pollen, spores, yedoma, ice wedge, pollen spectra, paleorecostructions, vegetation, long distance transported pollen, verification, total annual thawing index
Введение
В последнее время палеотемпературные реконструкции на основании палинологических данных становятся все более продвинутыми и требуют все более сложного математического аппарата. Поскольку ископаемая пыльца использовалась в качестве палеоэкологического и палеоклиматологического показателя в течение многих десятилетий [1], интенсивные исследования в этом направлении привели к появлению большого количества работ с оценкой сильных и слабых сторон интерпретации ископаемых палиноспектров в качестве палеоклиматического индикатора [1-8]. Развивающееся моделирование палеоклиматических условий демонстрирует существенные расхождения для позднего плейстоцена [9]. Некоторые из этих расхождений могут быть вызваны тем фактом, что многие палеоклиматические данные отражают определенный сезон, в то время как сигнал часто интерпретируется как среднегодовое значение [10], или потому, что даже в регионах, расположенных близко друг к другу, регистрируются существенные климатические градиенты. Часто при моделировании к данным по палеорастительности подходят со значительным осреднением, что не способствует точности результатов моделирования, в частности это касается летних температур.
Данные палинологического анализа могут быть использованы как для реконструкций растительного покрова, так и для получения информации о палеоклиматических показателях, поскольку базовое положение палинологического анализа состоит в том, что, при тех или иных определённых эдафических условиях и отсутствии вмешательства человека, климатические условия для каждой единицы растительности и ее пространственный предел соответствуют пороговым значениям. Растительность Арктики подходит для интерпретации в терминах палеотемпературы как нельзя лучше. На изменения климата наиболее резко реагирует растительность, произрастающая в краевых частях ареалов. Изучение изменения таких границ, помогает оценить изменения растительного покрова и температур периодов вегетации данной территории.
Для арктических территорий важно определить непосредственно границу Арктики. Для интерпретации данных палинологического анализа как представляется автору, больше всего подходит определение этой границы у А.И. Толмачева [11], который считает границей Арктики северную границу непрерывного распространения редколесий (рис. 1).
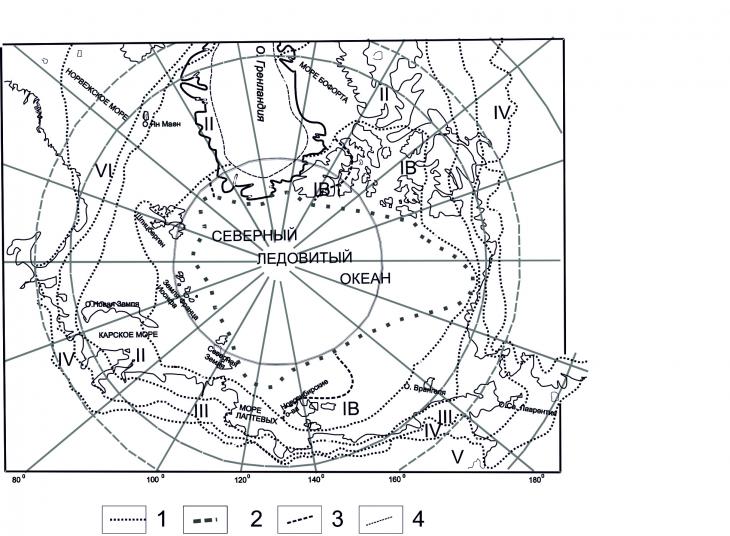
Рис. 1. Граница растительности Арктики по А.И.Толмачеву [11]: I – подзона высокоарктических тундр, IА – приледниковый гумидный вариант, IБ – приледниковый криоаридный вариант, I – внеледниковый вариант, II – подзона арктических тундр, III – подзона северных гипоарктических тундр, IV – подзона южных гипоарктических тундр, V – подзона крупных стлаников, VI – океанические аналоги подзон IV и V за пределами собственно тундровой зоны; 1 – границы подзон, 2 – границы абиссальных глубин Арктического бассейна, 3 – границы вариантов подзоны высокоарктических тундр, 4 – граница Гренландского ледникового щита
Севернее этой границы арктические и аркто-альпийские виды составляют значительную часть флоры, а южнее они играют незначительную роль [11]. В область Арктики также им включены: области крупных стлаников в бассейнах Анадыря и Пенжины, северная окраина Скандинавии, Исландия, Гренландия. Но здесь в сложении флоры аномально высокая роль принадлежит бореальным, океаническим гипоарктическим альпийским и аркто-альпийским видам (сформированные здесь субфоссильные палиноспектры практически неотличимы от спектров северной тайги). Широкое признание такой границы обусловлено тем, что она тесно связана с важнейшим геоботаническим рубежом – южной границей зоны тундр, или северной границей древесной растительности. На положение северного предела леса воздействует вторжение холодных арктических вод и льдов. Например, длительное стояние льдов в Гудзоновом заливе обусловливает расположение северной границы леса около 50° с.ш. Из-за этого различие между самым южным и самым северным расположением границы леса в северном полушарии составляет 20°. В Евразии северная граница леса, проходя параллельно береговой линии, колеблется от 72°30΄с.ш. в низовьях Лены до 58°55΄с.ш. на побережье Охотского моря. Именно эта граница может быть определена палинологически, как граница существенного падения концентрации пыльцы древесных пород на фоне высокой концентрации пыльцы типичных тундровых таксонов (Artemisia, Ericaceae, Poaceae, Betulasect. Nanae и др.), за исключением области крупных стлаников. На наш взгляд районирование растительного покрова Арктики по А.И.Толмачеву [11] лучше всего подходит для определения аналогов при палеореконструкциях, хотя и требует дополнительных данных, в частности, характеристик температуры и влажности, как основных факторов.
Для количественной оценки летних температур и фенологических связей удобно использовать показатель суммы среднесуточных положительных температур воздуха. Для растений средней полосы в качестве показателя используется сумма эффективных температур (сумма температур выше или равных +5⁰С). Для начала продуцирования пыльцы лиственных пород (Populus, Betula, Salix, Acer, Ulmus,) эта сумма составляет 70–80 ⁰С, исключение – Tilia (645– 690 ⁰С), хвойных (Picea, Pinus) – 180–200 ⁰С, – Poaceae – 400 ⁰С, а, например, для Tussilago farfara L. (мать-и-мачехи) всего +5.3⁰С, т.е. всего один день со средней температурой выше 5⁰С [12]. Продолжительность вегетационного периода зависит главным образом от географической широты и климатических условий. Вегетационный период - это время, необходимое для развития растения: для однолетнего - от прорастания семян до созревания семени; для многолетнего - от всхода семян или набухания почек до созревания семени; для деревьев и кустарников от начала сокодвижения и распускания почек до опадания листьев. Тундровые растения в основном многолетники, поскольку это позволяет в кратчайшие сроки переходить к активному росту. Для растений тундровой зоны таким показателем, объединяющим длительность сезона вегетации и температурный режим, является сумма положительных температур, поскольку, например, в пределах арктических тундр среднесуточные температуры могут не превышать +5⁰. Эта характеристика является нормативной, она приводится во всех климатических справочниках. Как оказалось, связь между составом палиноспектров и суммами среднесуточных положительных температур воздуха имеет детерминированный характер, хотя выразить это строго математически довольно сложно (рис. 2). На основании позднеплейстоценовых палинореконструкций именно этот показатель можно оценить достаточно адекватно.
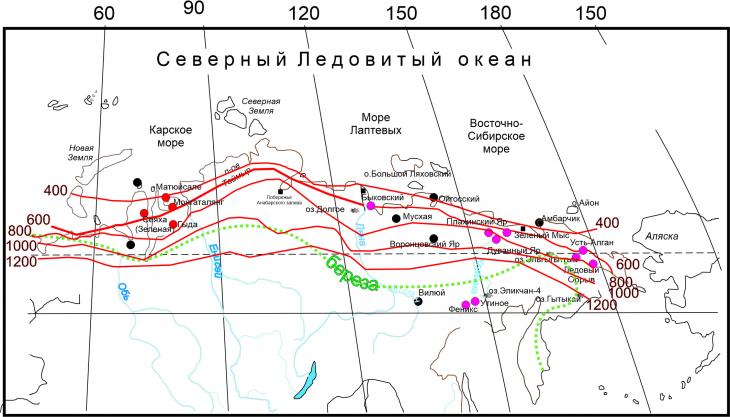
Рис. 2. Изотермы суммы среднесуточных положительных температур для криолитозоны России [13].
Особенно это проявляется при районировании тундровой зоны, что особенно важно для позднеплейстоценовых реконструкций растительности и климата. Ограничением данного метода является его точность, что связано с воздействием на процесс формирования палиноспектров ряда тафономических факторов.
В настоящее время реконструкции по палинологическим данным производятся в основном с использованием понятия биом, который является ключевой экологической и биогеографической единицей. Введение понятия биома привело к смещению акцентов при моделировании растительности прошлых эпох акцента с описания чистых моделей на понимание функциональности растительного покрова. Это также сформировало современное понимание концепции биома как динамической биологической сущности, которая имеет глубокие корни в эволюционном прошлом и претерпевающая изменения в настоящем [14, 15, 2].
Разделение спорово-пыльцевых диаграмм из Арктики на информационные блоки лучше всего проработано в проекте САРЕ (Circum Polar Arctic Environment - циркумполярная арктическая среда обитания), участники которого подготовили специальную процедуру биомизации для интерпретации палинологических данных в терминах палеотемператур [16]. Под биомизацией подразумевается объединение палиноспектров в группы согласно сочетаниям функциональных типов растений [17]. Собственно биомизация палинологических данных - это процесс, посредством которого предполагаемая растительность может быть представлена относительно небольшим числом биомов, описывающих основные структурные, функциональные и флористические различия. Для процедуры биомизации в модели BIOME4 отобрано ограниченное количество функциональных типов растений. Для интерпретации палинологических данных севера Евразии мы использовали классификацию биомов [18] и сопоставление состава палиноспектров с функциональными типами растений по Н. Биджелоу с соавторами [19] (табл. 1). Очевидно, что поскольку выделенные биомы не отражают всех компонентов палиноспектра, то коррекция значения суммы температур производилась с учетом состава конкретного палиноспектра выделенной палинозоны, с учетом классификации растительного покрова Арктики, предложенной А.И.Толмачевым [11].
Таблица 1. Классификация циркумполярных биомов (по Kaplan et al. [18])
|
Биом
|
Определение
|
Типичные таксоны
|
|
Низко- и высоко-
кустарниковая
тундра
|
Cомкнутый кустарниковый покров, кустарники высотой от 50 см до 2 м, лиственные или вечнозеленые иногда с образующими кочки граминоидами и с зелеными мхами, сфагновыми мхами и лишайниками
|
Alnus, Betula, Salix, Pinuspumila (в Восточной Сибири), Eriophorum, Sphagnum
|
|
Высококустарниковая
тундра
|
Cомкнутый кустарниковый покров, кустарники высотой от 2 см до 50 см, лиственные или вечнозеленые с граминоидами, с зелеными мхами и лишайниками
|
Betula, Cassiope, Empetrum, Salix, Vaccinium, Gramineae, Cyperaceae
|
|
Кустарничковая тундра
|
Несомкнутый растительный покров стелющихся лиственных кустарников высотой 0–2 см
|
Salix,Dryas, Pedicularis, Asteraceae, Caryophyllaceae, Gramineae, Bryales
|
|
Моховая –лишайниковая тундра с подушковидными травами
|
Несомкнутый растительный покров розеточные или подушковидные травы с лишайниками и мхами
|
Papaver, Draba, Saxifragaceae, Caryophyllaceae, lichens, Bryales
|
|
Граминоидная тундра с травами
|
Преимущественно травянистая растительность с граминоидами зелеными мхами и лишайниками.
|
Artemisia, Kobresia, Brassicaceae, Asteraceae, Caryophyllaceae, Gramineae, Bryales
|
Эта простая классификация позволяет выделить реакцию растительности на изменения летней температуры и влажности.
Таблица 2. Сопоставление классификации арктических биомов [16, 18,19], палинологическая характеристика выделенных биомов, соответствующие суммы летних температур (по данным авторов)
|
Растительная
подзона
|
Характер растительности и палиноспектров
|
Характеристика: длительность вегетационного периода, сумма положительных температур.
|
|
Полярные пустыни и тундры высоко- арктические
|
Несомкнутый растительный покров
|
Кустарнички отсутствуют
|
Преобладают споры Selaginella или Bryales,
заметное участие дальнезано-сной пыльцы деревьев
|
Наиболее низкие темпе-ратуры вегета-
ционного пери-
ода
|
30-45 дней
50-150 град. х дней
|
|
Доминирует пыльца трав, часто пыльцевые зерна недораз-виты,
|
Низкие темпе-ратуры вегета-ционного периода
|
30-45 дней
100-300 град. х дней
|
|
Кустарнички присутствуют
|
Наряду с пыльцой трав и спорами присутствует пыльца Salix
в качестве региональных компонетов пыльца Betula sect. Nanae и Alnaster,
|
Низкие темпе-ратуры
вегетацион-ного
периода
|
45-60 дней
200-400 град х дней
|
|
Гипоаркти-ческие тундры
|
Злаковые тундры
|
Преобладание злаков
|
Доминирование пыльцы Poaceae
|
Средние температуры вегетационного
периода
|
50-60 дней
350-700 град. х дней
|
|
Осоково-злаковые тундры
|
Домини-рование Poaceae + Cyperaceae
|
|
Злаково-полынные тундры с разно-травными групппиров-ками
|
Доминирование пыльцы Poaceae и Artemisia, с участием разнотравья
|
рост темпе-ратур вегетацион-ного периода
|
|
Вересковые тундры
|
Преоб-ладание вереско-цветных
|
Пребладание пыльцы Ericaceae
|
50-60 дней
450-750 град. х дней
|
|
Кустар-никово-злаковые
тундры
|
Ерниково-злаковые ас-социации
|
Преобладание пыльцы Betula sect. Nanae, Poaceae
|
рост темпе-ратур вегета-ционного пе-риода
|
50-60 дней
500-800 град.х дней
|
|
Кустар-никовые тундры
|
Ассоциации ерника и ольховника
|
Доминирование пыльцы Betula sect. Nanae и/или Alnaster
|
рост темпе-ратур вегета-ционного пе-риода
|
|
Крупно-кустар-никовые тундры
|
Ассоциации с кедровым стланником
|
Доминирование пыльцы Pinuspumila
|
рост темпе-ратур вегета-ционного пе-риода
|
50-60 дней
700-1200 град. х дней
|
|
Лесотундра
|
Лесотундровые формации
|
Тундровые ассоциации + лиственница
Тундровые ассоциации +
береза
Тундра +чозения
|
Доминирование пыльцы Betula sect Albae с участием пыльцы Larix
или хвойных, но с элементами тундровых палиноспектров
|
рост темпе-ратур вегетацион-ного пери-ода
|
|
Лес
|
Таежные формации
|
Домини-рование листвен-ницы,
березы
ели
сосен
|
Почти полное доминирование пыльцы
древесных видов
|
Рост темпе-ратур вегета-ционного периода
|
Более 60 дней,
выше 1200-1300 град.хдней
|
Наши исследования по изучению соотношения длительности вегетационного сезона, а именно изучение массива субфоссильных спектров из ряда регионов Арктики [20,21], позволили включить в характеристику соотношения биомов и палиноспектров сумму положительных температур в качестве синтетического показателя, отражающего и средние летние температуры и длительность летнего сезона (табл. 2). Сумма эффективных температур менее удобна, поскольку для некоторых районов температура летнего сезона не поднималась выше +5°С.
Применение данной методики реконструкции суммы положительных температур позволяет определить относительные изменения палиноспектров в отдельных образцах и выразить их в единых терминах суммы положительных температур. Следовательно, предложенная интерпретация позволяет сопоставлять между собой датированные спорово-пыльцевые диаграммы разрезов, расположенных на значительном удалении друг от друга. Разумеется, реконструкции климатических показателей с помощью переходных функций и статистических показателей [21-24], учитывающих соотношение каждого палинотаксона с температурами июля и среднегодовыми температурами позволяет реконструировать, палеотемпературы более точно, особенно для голоцена. Преимущество предложенной здесь методики реконструкций сумм положительных температур позволяет точнее интерпретировать опубликованные ранее спорово-пыльцевые диаграммы по разрезам едомных толщ, по которым информация представлена исключительно в графическом виде, и отследить резкие изменения в составе палиноспектров интерпретируя их в терминах суммы положительных температур (рис. 3).
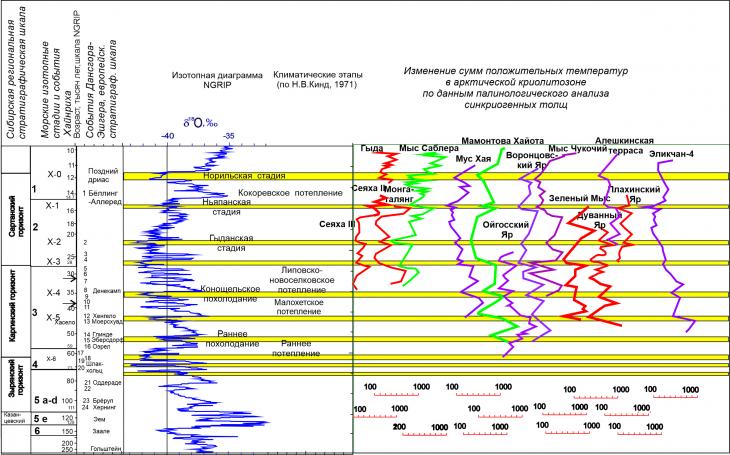
Рис. 3. Корреляция палеотемпературных кривых сумм положительных температур, полученных в результате авторской интерпретации датированных по 14С спорово-пыльцевых диаграмм по разрезам: Мыс Саблера - А.А.Андреева и Г.Н.Шиловой (Andreev et al. [25,26]); Мус-Хая Н.О.Рыбаковой и Л.Л.Козачихиной [27], Томирдиаро, Чёрненький [28]); Мамонтова Хайота - А.А.Андреева [29]; Воронцовский Яр - Н.С.Болиховской [30]; по Ойгосский Яр и Мыс Чукочий - Л.Л. Козачихиной [28]; по Алёшкинской террасе - И.Н. Федоровой и А.В. Ложкина [28]; по Эликчан-4 - А.В.Ложкина, П.Андерсон и Л.Брубейкер [31]; по разрезам Гыда, Сеяха-II и III, Монгаталянг, Зелёный Мыс, Плахинский Яр и Дуванный Яр – А.К. Васильчук [31-36]. Колебания содержания изотопов кислорода в керне NorthGRIP и привязка событий Хайнриха и Дансгора – Эшгера, согласно данным В.Дансгора, Х.Хайнриха, Д.Даль-Енсен, Дж. Бонда, Дж. Доудсвелла, Л.Хиннов [37, 38, 39, 40, 41, 42]
В качестве стратотипа мы использовали изменение изотопного состава ледникового керна NGRIP Гренландского ледникового щита, интерпретируемое как в большой мере обусловленное изменением среднегодовых температур. Вполне естественно предположить, что глобальные события отражались и на составе растительного покрова Северной Евразии и, следовательно, могут быть запечатлены на спорово-пыльцевых диаграммах. Изучение разрезов криолитозоны с учетом особенностей формирования многолетнемерзлых отложений позволило привязать имеющиеся данные к ритмам глобальных изменений среднего масштаба – событиям Хайнриха (см. рис. 2). Сопоставление кривых сумм положительных температур между собой выявило ряд общих трендов. Во-первых отмечается близкое сходство полученных распределений на региональном уровне. Так, кривые сумм положительных температур Сеяха 3 и Монгаталянг совпадают почти по всем ключевым точкам, как минимумам, так и максимумам. Совпадения по максимумам и минимумам отмечается также для более удаленных друг от друга разрезов: группы западно-сибирских разрезов - Сеяха 3 и Сеяха 2 Монгаталянг- и таймырского Мыс Саблера. Следовательно, можно говорить, что условия летних сезонов в данном временном интервале изменялись практически синхронно на севере Западной Сибири и Таймыре. Палеотемпературные кривые Зеленого Мыса, Дуванного Яра и Плахинского Яра также имеют ряд общих черт. Сходство между кривыми, полученными по данным из разрезов в долине р.Колымы выше, чем с кривыми полученными в едомных разрезах на севере Западной Сибири и Таймыра. Отметим очень близкую палинологическую характеристику разрезов Плахинского Яра и Алешкинской террасы и практически полное совпадение на одинаково датированных фрагментах колебаний сумм положительных температур.
Имеются также общие моменты, зафиксированные почти на всех кривых сумм положительных температур из проинтерпретированных спорово-пыльцевых диаграмм датированных по радиоуглероду разрезов синкриогенных отложений с повторно-жильными льдами. На полученных кривых заметны минимумы, соответствующие похолоданиям, связанным с событиями Хайнриха, это означает, что в эти периоды летние сезоны отличались низкими температурами, а их длительность также снижалась (рис.4). Сумма положительных температур для этих периодов не превышала 300 град. х сут. Распределение сумм положительных температур для стадиалов событий Хайнриха практически не отличалось. В то время как интерстадиальный растительный покров различался по доминирующим компонентам и распределение сумм положительных температур для каждого периода летнего потепления было более индивидуальным.
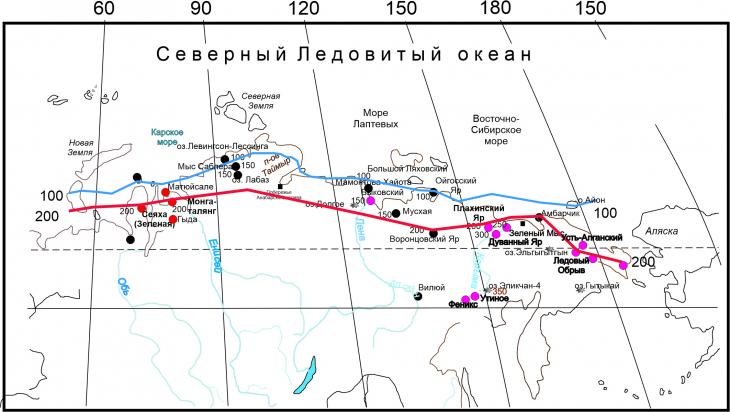
Рис. 4. Распределение значений сумм положительных температур в минимальную (стадиальную) фазу третьего события Хайнриха 28-27 т.л.н.
Возможно, снижение температур вегетационного периода было связано с увеличением площади ледового покрова в летние сезоны. Например, выделяется минимум летних температур, соответствующий третьему событию Хайнриха примерно 28,5-27 тыс. лет назад. Интересно, что, наиболее заметное увеличение сумм положительных температур, за которым следует резкое их снижение отмечается в начале каждого события Хайнриха.
Исследованиями М Стайвера и П.Грутса [43] установлено, что серджи, связанные с событиями Хайнриха, происходили в течение холодного пика отдельных событий Дансгора-Эшгера, выделяемых на изотопных диаграммах гренландских ледниковых кернов, средняя продолжительность цикла Дансгора–Эшгера составляла около 1460 лет, при этом холодная фаза длилась около 600 лет и заканчивалась резким переходом к теплой фазе. То есть, судя по реакции растительного покрова в Северной Евразии и данным изотопного состава ледяных жил [44], наиболее выраженным было летнее потепление, которое прерывалось резким похолоданием, связанным с поступлением айсбергового материала в моря Северного Ледовитого океана.
Общий позитивный пик отмечен около 29 тыс. лет практически на всех кривых положительных температур. Он соответствует интерстадиалу третьего события Хайнриха. На кривых Сеяха-2 и Зеленый Мыс можно выделить существенное снижение температур вегетационного периода, которое может соответствовать первому событию Хайнриха, а затем последующий резкий рост, который может быть сопоставлен с аллередом. Это потепление на севере Западной Сибири проявилось несколько ярче с большей амплитудой. На кривой Гыда удалось выделить период снижения сумм положительных температур, сопоставляемый с поздним дриасом, и последующий переход к гренландскому периоду голоцена. Можно утверждать, что глобальные климатические колебания, обусловленные изменениями ледовой обстановки в Северной Атлантике – событиями Хайнриха особенно заметно отражались на растительном покрове современной криолитозоны, т.е климатические характеристики летнего сезона изменялись весьма существенно. Разумеется, каждая наземная система по-разному реагирует на климатические изменения, что приводит к различному времени отклика и усложняет хронологию. По результатам проведения конференции «События Хайнриха в наземных архивах” на конгрессе INQUA в Дублине в 2019 году, события Хайнриха отразились на составе ряда наземных объектов: эоловых отложений [45], в Ауэль-Маара в Германии, на составе растительности и остракод в оз. Петен-Ица в Гватемале, а также на изотопном составе спелеотем. С. Вететрих и Н. Рудая с [46], на основании изучения пыльцы и спор в разрезе Собо-Сисе и сопоставления их с синхронно формировавшимися едомами пришли к заключению, что региональные пыльцевые записи в дельте Лены и на Быковском полуострове подтверждают существование позднеплейстоценовой тундро-степи в Западной Берингии во время MIS 3 с частично более теплым, чем сегодня, летом и более влажными условиями. Наиболее благоприятные условия отмечаются для интерстадиала после похолодания пятого события Хайнриха, что совпадает с максимумом суммы положительных температур для кривой Зеленый Мыс (700-750 град.х дней.). Согласно их выводам С Веттериха [46], во время MIS 2 лето, вероятно, было более холодным, чем в течение MIS 3, и более сухим с нестабильными условиями увлажнения в полигональных ландшафтах. Согласно реконструкциям суммы положительных температур (рис. 2) в течение интерстадиалов 2, 3 и 4 событий Дансгора-Эшгера (стадии MIS2) сумма положительных температур была на 100-150 град.хдней ниже чем в течение интерстадиалов с 5 по 15 событий Дансгора –Эшгера.
По результатам реконструкций состава палиноспектров из отложений оз. Лирней [47] минимальное значение среднеиюльской температуры 7.7°C установлено 27.90 тыс. кал. лет назад, а максимальное 9.3°C, 15.72–15.47 тыс. кал. лет назад. На основании результатов моделирования с использованием данных палинологического анализа и изменений термогалинной циркуляции поверхностных и глубинных вод Северной Атлантики [48] получен вывод, что интерстадиалы событий Дансгора-Эшгера в Западной Европе характеризовались вдвое большей суммой осадков по сравнению со стадиалами, температура самого холодного месяца была на 17°C выше, а самого теплого месяца на 8°C выше, реконструированная сумма положительных температур составила около 1300 °C х дней. Cовокупный эффект низких температур в зимний период, недостаточное увлажнение летом и низкое значение суммы эффективных температур, полученное по результатам моделирования, объясняет по мнению авторов отсутствие лесов к северу от 48°с.ш. во время межстадиала 14 события Дансгора-Эшгера [48] несмотря на относительно благоприятные условия вегетационного сезона. На наш взгляд, причиной этому не недостаточная сумма эффективных температур, а прежде всего короткий сезон вегетации. Поэтому в разрезах позднеплейстоценовой палеокриолитозоны в Европе встречается пыльца травянистых растений, требовательных к тепловому режиму и в настоящее время произрастающих в лесной зоне [49], а пыльца древесных пород отмечается в небольших количествах, не соответствующих лесным формациям. По мнению авторов данной статьи, при построении моделей для реконструкции растительного покрова и температуры сезона вегетации и длительность сезона вегетации или не учитываются, или учитываются недостаточно.
В обзорной статье М.Шевалье с соавторами [2], дана оценка 75 летнему опыту разработки и применения количественных методов реконструкции климата на основании палинологических данных, начиная с пионерных реконструкции Й. Иверсена [50]. Несмотря на успешную разработку альтернативных климатических индикаторов в последние годы, в том числе основанных на изотопах и биомаркерах, ископаемая пыльца, по мнению авторов обзора [2], остается наиболее важным источником количественных климатических данных для наземных неледниковых регионов, например [51]. С развитием новейших статистических способов интерпретации, количественные реконструкции стали намного доступнее.
Выводы
Предложена оригинальная методика реконструкции суммы положительных температур, основанная на соотношении биомов и палиноспектров в качестве объективного показателя, отражающего средние летние температуры и длительность летнего сезона. Это позволяет оценить относительные изменения палиноспектров в отдельных образцах и выразить их в единых терминах суммы положительных температур воздуха.
Интерпретация палиноспектров, как отражение суммы положительных температур позволила сопоставить между собой датированные спорово-пыльцевые диаграммы разрезов, расположенных на значительном удалении друг от друга.
Сопоставление кривых сумм летних температур воздуха между собой выявило ряд общих трендов: а) близкое сходство полученных распределений на региональном уровне; б) синхронность изменений условий летних сезонов на севере Западной Сибири и Таймыре; в) сходство диаграмм сумм летних температур воздуха, полученных по данным опорных позднеплейстоценовых разрезов в долине р. Колымы.
Установлено хронологическое соответствие минимумов сумм летних температур воздуха на полученных диаграммах и событий Хайнриха.
References
1. Birks, H.J.B., 2019. Contributions of Quaternary botany to modern ecology and biogeography. Plant Ecol. Divers. 12, 189–385. https://doi.org/10.1080/17550874.2019.1646831.
2. Chevalier M., Davis B.A.S., Oliver Heiri B. et al. Pollen-based climate reconstruction techniques for late Quaternary studies // Earth-Science Reviews. 2020. Vol. 210, 103384. doi: 10.1016/j.earscirev.2020.103384.
3. Birks, H.J.B., 2014. Quantitative palaeoenvironmental reconstructions from Holocene biological data. Glob. Chang. Holocene 9780203785, 107–123. https://doi.org/10. 4324/9780203785027.
4. Vasil'chuk A.K. Osobennosti formirovaniya palinospektrov v kriolitozone Rossii. M.: Izd-vo Mosk. un-ta, 2005. 245 s.
5. Vasil'chuk A.K. Palinologiya i khronologiya poligonal'no-zhil'nykh kompleksov v kriolitozone Rossii. M.: Izd-vo Mosk. un-ta, 2007. 488 s.
6. Gajewski, K., 1993. The role of paleoecology in the study of global climatic change. Rev. Palaeobot. Palynol. 79, 141–151. https://doi.org/10.1016/0034-6667(93)90044-U.
7. Gajewski, K., 2015. Quantitative reconstruction of Holocene temperatures across the Canadian Arctic and Greenland. Glob. Planet. Change 128, 14–23. https://doi.org/ 10.1016/j.gloplacha.2015.02.003.
8. Webb, T.I., 1986. Is vegetation in equilibrium with climate? How to interpret Late-Quaternary pollen data. Vegetation 67, 75–91. https://doi.org/10.1007/BF00037359.
9. Velasquez P., Kaplan J.O., Messmer M., Ludwig P., Raible C.C. The role of land cover in the climate of glacial Europe Clim. Past, 17, 1161–1180, 2021 https://doi.org/10.5194/cp-17-1161-2021
10. Beghin, P., Charbit, S., Kageyama, M., Combourieu-Nebout, N., Hatté, C., Dumas, C., and Peterschmitt, J.-Y.: What drives LGM precipitation over the western Mediterranean? A study focused on the Iberian Peninsula and northern Morocco, Clim. Dynam., 46, 2611–2631, https://doi.org/10.1007/s00382-015-2720- 0, 2016
11. Tolmachev A.I. Vvedenie v geografiyu rastenii L.: LGU, 1974, 244 s
12. Kolbina L.M., Osokina A.S Summa effektivnykh temperatur i raboty na paseke v Udmurtskoi Respublike Biomika, 2019 tom 11, № 2, s. 143-146 DOI
13. Spravochnik po klimatu SSSR. L. Gidrometeoizdat. Vo vsekh vypuskakh - Chast' 2. Temperatura vozdukha i pochvy. Vyp. 1.Arkhangel'skaya i Vologodskaya oblasti, Karel'skaya i Komi ASSR. 1965. 359 s. Vyp. 17. Tyumenskaya i Omchkaya oblasti. 1965. 400 s. Vyp. 20. Tomskaya, Novosibirskaya, Kemerovskaya oblasti i Altaiskii krai. 1966. 396 s. Vyp. 21. Krasnoyarskii krai i Tuvinskaya ASSR. 1966. 504 s. Vyp. 23. Buryatskaya ASSR i Chitinskaya oblast'. 1966. 319 s. Vyp. 24. Yakutskaya ASSR. 1966. 403 s. Vyp. 33. Chukotskii natsional'nyi okrug i Magadanskaya oblast'. 1967. 288 s.
14. Peyron, O., Guiot, J., Cheddadi, R., Tarasov, P.E., Reille, M., de Beaulieu, J.-L., Bottema,S., Andrieu, V., 1998. Climatic reconstruction in Europe for 18,000 YR B.P. from pollen data. Quat. Res. 196, 183–196. https://doi.org/10.1006/qres.1997.1961.
15. Bartlein, P.J., Harrison, S.P., Brewer, S.C., Connor, S., Davis, B.A.S., Gajewski, K., Guiot,J., Harrison-Prentice, T.I., Henderson, A., Peyron, O., Prentice, I.C., Scholze, M.,Seppa, H., Shuman, B.N., Sugita, S., Thompson, R.S., Viau, A.E., Williams, J.W., Wu,H., 2011. Pollen-based continental climate reconstructions at 6 and 21 ka: a global synthesis. Clim. Dyn. 37, 775–802. https://doi.org/10.1007/s00382-010-0904-1.
16. CAPE Project members. Holocene paleoclimate data from the Arctic: testing models of global climate change // Quaternaty Sciences Reviews. 2001. Vol. 20. P.1275 – 1287.
17. Prentice, I.C., Guiot, J., Huntley, B., Jolly, D., Cheddadi, R., 1996. Reconstructing biomes from palaeoecological data: a general method and its application to European pollen data at 0 and 6 ka. Clim. Dyn. 12, 185–194. https://doi.org/10.1007/BF00211617
18. Bigelow N.H., Brubaker L.B., Edwards M.E., Harrison S.M., Prentice I.C., Andreson P.M., Andreev A.A., Baetlein P.J., Christensen T.R., Cramer W., Kaplan J.O., Lozhkin A.V., Matveyeva N.V., Murray D.F., McGuire A.D., Razzhivin V.Y., Ritchie J.C., Smith B., Walker D.A., Gajewski K., Wolf V., Holmqvist B.H., Igarashi Y., Kremenetskii K., Paus A., Pisaric M.F.J., Volkova V.S. Climate change and Arctic ecosystems: I: Vegetation changes north of 55oN between the last glacial maximum, mid-Holocene, and present // Journal of geophysical research. 2003. Vol. 108. N D19. P. 8170 – 8194.
19. Kaplan, J.O., Bigelow, N.H., Prentice, I.C., Harrison, S.P., Bartlein, P.J., Christensen, T.R.,Cramer, W., Matveyeva, N.V., McGuire, A.D., Murray, D.F., Razzhivin, V.Y., Smith, B., Walker, D.A., Anderson, P.M., Andreev, A.A., Brubaker, L.B., Edwards, M.E., Lozhkin, A.V., Climate change and Arctic ecosystems: 2.Modeling, paleodata-model comparisons, and future projections. J. Geophys. Res. 2003. 108 (D19), 8171. http://dx.doi.org/10.1029/2002JD002559.
20. Vasil'chuk A.K. Regional'naya i dal'nezanosnaya pyl'tsa v tundrovykh palinospektrakh Izvestiya Rossiiskoi Akademii Nauk. Seriya biologicheskaya. 2005. №1. S. 85 – 99. Vasil'chuk A.K. 2005. Regional and extra-local pollen in tundra pollen samples // Biology Bulletin of the Russian Academy of Sciences. Vol. 32. N1. P. 75–84.
21. Vasil'chuk A.K. Osobennosti tafonomii pyl'tsy i spor v Arktike // Izvestiya Rossiiskoi Akademii Nauk. Seriya biologicheskaya. 2005. №2. S. 240–252. (Vasil'chuk A.K. 2005. Taphonomic features of Arctic pollen // Biology Bulletin of the Russian Academy of Sciences. Vol. 32. N2. P. 196–206).
22. Klimanov V.A. Klimat Severnoi Evrazii v pozdnelednikov'e i golotsene / Avtoreferat dissertatsii na soiskanie uchenoi stepeni doktora geograficheskikh nauk. M. IGAN. 1996. 46 s.
23. Izumi, K., Bartlein, P.J., North American paleoclimate reconstructions for the Last Glacial Maximum using an inverse modeling through iterative forward modeling approach applied to pollen data. Geophys. Res. Lett. 2016. 43, 10,965–10,972. https://doi. org/10.1002/2016GL070152.
24. Binney, H.A., Edwards, M.E., Macias-Fauria, M., Lozhkin, A., Anderson, P., Kaplan, J.O., Andreev, A.A., Bezrukova, E., Blyakharchuk, T.A., Jankovska, V., Khazina, I., Krivonogov, S., Kremenetski, K.V., Nield, J., Novenko, E.Y., Ryabogina, N., Solovieva, N., Willis, K.J., Zernitskaya, V.P., Jankovska, V., 2017. Vegetation of Eurasia from the last glacial maximum to present: key biogeographic patterns. Quat. Sci. Rev. 157,80–97. https://doi.org/10.1016/j.quascirev.2016.11.022.
25. Andreev A.A., Schirrmeister L., Siegert C., Bobrov A.A., Demske D., Seiffert M., Hubberten H.-W. Paleoenvironmental changes in Northeastern Siberia during the Late Quaternary - evidence from pollen records of the Bykovsky Peninsula // Polarforschung. 2002. Vol. 70. P. 13 – 25.
26. Andreev A.A., Tarasov P.E., Siegert Ch., Ebel T., Klimanov V.A., Melles M. , Bobrov A.A., Dereviagin A.Yu., Lubinski D.J., Hubberten H.-W. Vegetation and climate changes on the northern Taymyr, Russia during the Upper Pleistocene and Holocene reconstructed from pollen records // Boreas. 2003. Vol. 32. N3. P. 484 – 505.
27. Kondrat'eva K.A., Trush N.I., Chizhova N.I., Rybakova N.O. K kharakteristike pleistotsenovykh otlozhenii v obnazhenii Mus-Khaya na r.Yane // Merzlotnye issledovaniya, vyp. 15. M.: Izd-vo Mosk. un-ta. 1976. S. 60 – 93.
28. Tomirdiaro S.V., Chernen'kii B.I. Kriogenno-eolovye otlozheniya Vostochnoi Arktiki i Subarktiki. Otv. red. S.F.Biske. M.: Nauka, 1987. 198 s.
29. Schirrmeister L., Siegert C., Kuznetsova T., Kuzmina S., Andreev A., Kienast F., Meyer H., Bobrov A. Paleoenvironmental and paleoclimatic records from permafrost deposits in the Arctic region of Northern Siberia // Quaternary International. 2002. Vol. 89. Iss. 1. P. 97 – 118.
30. Bolikhovskaya N.S., Bolikhovskii V.F. Iskopaemye pochvy v lessovidnykh otlozheniyakh Severo-Vostoka Evrazii // Doklady AN SSSR. 1979. Tom 247. №2. S. 409 – 412.
31. Lozhkin A.V., Anderson P.M., Belaya B.V. Radiouglerodnoe datirovanie pyl'tsevykh zon v ozernykh osadkakh Kolymo-Okhotskogo mezhdurech'ya // Doklady Rossiiskoi Akademii Nauk. 1995. Tom 343. №3. S. 396 – 399.
32. Vasil'chuk A.K. Palinospektry 14S datirovannykh obraztsov sinkriogennykh porod severa Zapadnoi Sibiri i Yakutii // Kriosfera Zemli. 2002. Tom VI. №2. S. 3–21.
33. Vasil'chuk A.K. Otrazhenie sobytii Khainrikha na datirovannykh po radiouglerodu sporovo-pyl'tsevykh diagrammakh povtorno-zhil'nykh l'dov i vmeshchayushchikh ikh edomnykh otlozhenii nizovii r.Kolymy // Kriosfera Zemli. 2003. Tom VII. №4. S. 3–13.
34. Vasil'chuk A.K. Radiouglerodnoe datirovanie pyl'tsy i spor iz povtorno-zhil'nykh l'dov Yamala i Kolymy // Izvestiya Rossiiskoi Akademii Nauk. Seriya biologicheskaya. 2004. №2. S. 225 – 237. (Vasil'chuk A.K. 2004. Radiocarbon Dating of Pollen and Spores from Ice Wedges of the Yamal and Kolyma Regions // Biology Bulletin of the Russian Academy of Sciences. Vol. 31. N2. P. 182–192).
35. Vasil'chuk A.K. Palinospektry pozdnepleistotsenovykh i golotsenovykh povtorno-zhil'nykh l'dov v ust'e reki Seyakha na Vostochnom Yamale // Kriosfera Zemli. 2005g. Tom IX. №2. S. 43 – 53.
36. Vasil'chuk A.K. Korrelyatsiya palinospektrov iz pozdnepleistotsenovykh singeneticheskikh poligonal'no-zhil'nykh kompleksov // Materialy Tret'ei konferentsii geokriologov Rossii. MGU im. M.V.Lomonosova, 1 – 3 iyunya 2005 g. Moskva. Tom. 3. Chast' 6. Regional'naya i istoricheskaya geokriologiya. Izd-vo Mosk. un-ta. 2005e. S. 35 – 42.
37. Heinrich, H., 1988. Origin and consequences of cyclic ice rafting in the Northeast Atlantic Ocean during the past 130,000 years. Quaternary Research 29, 142–152.
38. Dansgaard, W. et al. A new Greenland deep ice core. Science. 1982. 218, 1273–1277.
39. }Bond, G.C., Broecker, W.S., Johnsen, S., McManus, J.F., Labeyrie, L., Jouzel, J., Bonani, G., 1993. Correlation between climate records from North Atlantic sediments and Greenland ice. Nature 365, 143–147.
40. Dowdeswell J.A., Maslin M.A., Andrews J.T., McCave I.N. Iceberg production, debris rafting, and the extent and thickness of Heinrich layers (H-1, H-2) in North Atlantic sediments // Geology. 1995. Vol. 23. P. 301 – 304.
41. Hinnov L.A., Schulz M., Yiou P. Interhemispheric space-time attributes of the Dansgaard-Oeschger oscillations between 100-0 ka // Quaternary Science Reviews. 2002. Vol. 21. P. 1213 – 1228.
42. Dahl-Jensen, D. Past temperatures directly from the Greenland Ice Sheet. Science. 1998. 282, 268–271
43. Stuiver, M., & Grootes, P. (2000). GISP2 Oxygen Isotope Ratios. Quaternary Research, 53(3), 277-284. doi:10.1006/qres.2000.2127
44. Vasil'chuk Yu.K., Kim J.-Ch., van der Plicht J., Papesch W., Vasil'chuk A.C. AMS-dating Dansgaard-Oeschger events in Siberian ice-wedge ice // 10th International Conference on Accelerator Mass Spectrometry. AMS-10. Conference Agenda & Abstracts. Berkeley, Calofornia, September, 5-10, 2005. P. 35 – 36
45. Fuhrmann, F., Seelos, K., & Sirocko, F. (2021). Eolian sedimentation in central European Auel dry maar from 60 to 13 ka. Quaternary Research, 101, 4-12. doi:10.1017/qua.2020.81
46. Wetterich S., Rudaya N., Nazarova L., Syrykh L., Pavlova M., Palagushkin O., Kizyakov A., Wolter J., Kuznetsova T., Aksenov A., Stoof-Leichsenring K. R., Schirrmeister L., Fritz M. Paleo-Ecology of the Yedoma Ice Complex on Sobo-Sise Island (Eastern Lena Delta, Siberian Arctic)// Frontiers in Earth Science. 2021. Vol. 9. 681511. doi: 10.3389/feart.2021.681511.
47. Andreev A. A., Raschke E., Biskaborn B. K., Vyse S. A., Courtin J., Bohmer T., Stoof-Leichsenring K., Kruse S., Pestryakova L. A., Herzschuh U. Late Pleistocene to Holocene vegetation and climate changes in northwestern Chukotka (Far East Russia) deduced fromlakes Ilirney and Rauchuagytgyn pollen records // Boreas, 2021.Vol. 50, pp. 652–670. doi: 10.1111/bor.12521.
48. Van Meerbeeck C.J., Renssen H., Roche D.M., Wohlfarth B., Bohncke S.J.P., Bos J.A.A., Engels S., Helmens K.F., Sánchez-Goñi M.F., Svensson A., Vandenberghe J.,The nature of MIS 3 stadial–interstadial transitions in Europe: New insights from model–data comparisons, Quaternary Science Reviews, 2011,Vol. 30, Iss. 25–26, P. 3618-3637/https://doi.org/10.1016/j.quascirev.2011.08.002.
49. Isarin R.F.B., Bohncke S.J.P. Mean July temperatures during the Younger Dryas in Northern and Central Europe as inferred from Climate Indicator Plant Species. Quaternary Research 1999. Vol. 51.158-173
50. Iversen J. Viscum, Hedera and Ilex as Climate Indicators. A Contribution to the Study of the Post-Glacial Temperature Climate. // Geologiska Föreningen i Stockholm Förhandlingar. 1944. Vol. 66. Iss. 3. P. 463–483. doi: 10.1080/11035894409445689.
51. Kaufman D., McKay, N., Routson, C. et al. A global database of Holocene paleotemperature records. Sci. Data. 2020 Vol. 7, 115 doi: 10.1038/s41597-020-0445-3.
|







